
0512-6856 7071
info@ecellbio.com
详谈TGF-β家族那些事
TGF-β家族介绍
TGF-β家族的信号蛋白广泛分布于动物中,其家族包括了TGF-β亚型、活化素(Activin)、抑制素(Inhibin)、骨形态发生蛋白(BMP)、生长分化因子(GDF)在内的40个已知成员组成。大致可分为TGF-β/Activin/Nodal和BMP/GDF/MIS (Muellerian inhibiting substrate)两个亚家族。这个家族的成员是细胞增殖、分化、机制合成、谱系分化、迁移、黏附和凋亡、胚胎发育、胞外基质形成、骨的形成和重建以及凋亡的关键调节者。它们在出生前发育和出生后生长、重塑和多种组织器官的维持中发挥主要作用。与它们调节重要生物过程的作用相一致,不同物种中同源TGF-β蛋白的氨基酸序列是高度保守的。该家族所有成员的共同特征有:(1)功能性配体是一个由两条12-15kDa称为单体的多肽链通过二硫键连接组成的同源或异源二聚体;(2)每个单体表达为前体肽的C-末端,同时包含分泌用的短的N-末端信号序列和一个长的200-300个氨基酸组成的长多肽;(3)单体初级结构的C-末端包含一个高度保守的七个半胱氨酸结构域。这些半胱氨酸残基中的一个用于形成链间二硫键桥,其他的参与形成分子内环的形成,即所谓的半胱氨酸结结构。半胱氨酸结是一个折叠的基序,可强制疏水残基暴露给周围的水,并阻止分子成为假定的球状蛋白结构(如图所示)。相反,它驱动分子发生二聚化,与一个蝶状结构形成一个高稳定性的二聚体蛋白。半胱氨酸结结构基序不仅在TGF-β蛋白中存在,也存在于其他半胱氨酸家族蛋白中,比如NGF、PDGF、糖蛋白激素(GPH)和IL-17等,因此组成了一个大的半胱氨酸结超家族。
TGF-β家族成员传导信号与涉及配体结合到I型和II型跨膜受体结构,诱导组成了一个活化的丝氨酸、苏氨酸激酶受体复合物。此受体复合物通过磷酸化胞浆中的Smad蛋白启始了一个信号转导途径,Smad蛋白磷酸化后转位于细胞核中,发挥抑制和激活靶基因转录的作用。应该指出的是,配体无障碍的接近TGF-β受体是受配体-或受体-结合蛋白高度控制的,配体-受体相互作用可被视为此信号途径的下游阶段,而不是上游。BAMBI(BMP和活化素膜结合蛋白)组成了通过充当BMP/活化素II型受体调节TGF-β信号的跨膜蛋白的一个范例。这个膜锚定蛋白与二型受体形成了稳定但信号不完全的复合物,因此抑制BMP/活化素信号。一个更广泛的调节类型涉及可溶性蛋白,与TGF-β配体形成潜在的复合物。除了为TGF-β配体提供细胞外储存位点,这些结合蛋白在控制TGF-β活性的局部浓度中发挥核心作用。TGF-β活性的缺陷与一系列临床适应症有关,包括肿瘤细胞生长、纤维化、骨骼缺陷和自身免疫性疾病等。因此,了解控制TGF-β信号的调节机制以及这些分子成分的识别和表征已成为当前研究的主要课题。
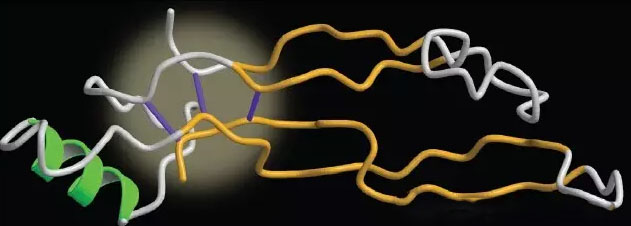
TGF-β家族半胱氨酸结构像的示意图,TGF-β家族半胱氨酸结单体结构由两个指状的反平行β链投影(金色),一个α-螺旋跟区域(绿色)。两个二硫键组成一个共价环,并被第三个二硫键渗透形成结状。
TGF-β亚型及其受体
在哺乳动物至少有TGF-β1、TGF-β2、TGF-β3及TGF-β1β2四个亚型。在鸟类和两栖类动物还分别存在着TGF-β4和TGF-β5,对后两者的生物学作用所知甚少。TGF家族蛋白结构相对比较保守,没有种属特异性,且其异构体活性基本相同。大多数正常细胞和肿瘤细胞都可分泌一种以上的TGF-β。单核巨噬细胞、中性粒细胞、成骨细胞、内皮细胞、淋巴细胞、成纤维细胞、巨核细胞、间皮细胞、软骨细胞和白血病细胞主要产生TGF-β1。角质细胞、卵泡粒膜细胞、神经胶质瘤细胞和猴肾 (BSC-1)等主要产生TGF-β2。间充质起源的细胞,如人黑色素瘤细胞(A-375)主要产生TGF-β3。提到细胞因子,我们总是会想到对应的受体。那么TGF-β的受体有哪些呢?TGF-βR存在着Ⅰ、Ⅱ、Ⅲ型三种形式,其主要信息可以见上表。TGF-βRⅠ和Ⅱ共同组成有活性的TGF-β受体,但两者参与不同的信号传递,且与不同的功能相关:TGF-β抑制细胞增值的作用必须有Ⅱ型受体存在;而Ⅰ型受体能介导TGF-β对细胞外基质的作用,并与TGF-β诱导的几种基因表达有关。
TGF-β的功能
1、抑制细胞增殖、炎症及免疫反应。
一方面TGF-β1可以调控细胞周期,使细胞停止在G1 期,或延长G1 期,进而抑制细胞增殖,诱导细胞分化或凋亡。
另一方面,TGF-β是有效的免疫抑制剂,可抑制机体的免疫功能,帮助肿瘤细胞逃避宿主免疫监视,并能抑制炎症。抑制免疫功能的TGF-β主要是TGF-β1,其它TGF-β可能主要作用于其它组织。TGF-β1对T 淋巴细胞和其他免疫活性细胞如自然杀伤细胞和单核细胞等起负性调节作用,还可拮抗数种白细胞介素及肿瘤坏死因子、干扰素的作用。
此外,在不同的细胞中可通过一个Smad3依赖的机制抑制主要组织相容性复合体Ⅱ类抗原转录激活物而抑制主要组织相容性复合体Ⅱ的表达。TGF-β1能够抑制多种T细胞活性:比如抑制T细胞表达细胞因子受体、抑制T细胞产生细胞因子和抑制T细胞的细胞毒性作用,它的抑制作用主要针对静止状态的T细胞,对已经活化的T细胞的作用未知。
2 、促进作用和趋化作用
TGF-β可促进间充质来源细胞的增殖和功能,促进施万(Schwann)细胞增殖,促进成骨细胞分裂和合成基质蛋白;刺激单核细胞分泌TNF、TGF-α、FGF、IL-1和TGF-β。TGF-β能诱导多种细胞趋化,如成纤维细胞、单核细胞、中性粒细胞及角质细胞等。
TGF-β还可以诱导多种细胞和分泌细胞外基质蛋白、促进产生蛋白水解酶抑制剂,并能促进细胞表达整合素受体,这对胚胎形成、骨组织和软组织愈合等有重要影响。
3 、TGF-β对肿瘤的作用
TGF-β1在肿瘤的发生、发展过程中表现为两种截然相反的作用:在肿瘤生长的早期, 肿瘤细胞表达少量的TGF-β1,TGF-β1可作为肿瘤的抑制物;但随着肿瘤的进展,肿瘤细胞较多地表达TGF-β1,此时TGF-β1成为促进肿瘤生长的物质。在肿瘤发生的后期,TGF-β1 直接作用于肿瘤细胞,加速肿瘤的进程。
TGF-β1加速肿瘤的进程,促进癌的侵袭和转移主要通过三方面的作用机制:增加瘤细胞与胞外基质的相互作用;增加血管生成;抑制免疫系统。
大量研究显示,TGF-β1和血管内皮生长因子的表达呈正相关。TGF-β1 刺激肿瘤血管生成的作用可通过旁分泌途径诱导血管内皮生长因子的表达,间接地刺激肿瘤血管的生成,同时血管内皮生长因子对 TGF-β1也有类似作用,二者共同促进血管的生成。
4 、 TGF-β调节炎症反应的作用
TGF-β是一种强大的免疫反应调节剂,它可以放大免疫反应,在特定的条件下更能有效地介导免疫反应。在炎症时,局部血小板释放的TGF-β使单核细胞和成纤维细胞趋化,进一步诱导单核细胞释放不同的生长因子和炎症介质,使炎症放大和损伤组织修复。
一旦单核细胞转化为巨噬细胞,TGF-β又能抑制巨噬细胞的呼吸爆发(IFN-γ和TNF-α能逆转这一作用),防止巨噬细胞的损伤作用,它还可以抑制TNF-α表达,抑制中性粒细胞和淋巴细胞与内皮细胞的粘附,从而起到抗炎的作用,阻止炎症对机体的伤害。
(如需TGF家族完整挂图请联系我们索取)



